LL-37: The Self-Defence Peptide That Also Turns on the Self
Humans carry a single cathelicidin. It kills microbes, calls in immune cells and helps wounds close — and, in the wrong context, teaches the immune system to attack its owner. A look at the most genuinely double-edged peptide in innate immunity.
LL-37 is the only human cathelicidin antimicrobial peptide. It kills microbes directly, neutralises bacterial LPS, recruits immune cells and supports wound healing. The same peptide also drives inflammation: in psoriasis, rosacea and lupus it converts self-DNA into an immune trigger. Any therapeutic use remains preclinical or early-stage and unproven.
One human gene encodes one cathelicidin, and from it comes one peptide — just 37 amino acids — that can sterilise a wound, summon white blood cells75, and, on a different day in a different tissue, teach the immune system to fire on its owner’s own DNA.15 Few molecules in innate immunity are so plainly two-faced, and fewer still are so honestly studied as both shield and saboteur.
What exactly is LL-37, and why only one?
Mammals carry a family of cathelicidins; humans carry exactly one, encoded by the CAMP gene and stored as an inactive precursor, hCAP18.7 Proteolytic cleavage — classically by proteinase 3 in neutrophils — releases the mature peptide7, named LL-37 for the two leucines at its start and its length.7 The fragment is cationic and amphipathic: a positively charged, helix-forming chain that is drawn to the anionic surfaces of bacterial membranes.7 That structural bias is the whole story of its first job. The peptide partitions into microbial membranes and disrupts them7, an action that does not depend on a single protein receptor and is therefore slow to provoke classical resistance.1
LL-37 is widely distributed — neutrophils, epithelial surfaces, skin, the airway — precisely the borders where host meets microbe.6 It is, in the literal sense, a peptide of frontiers.
How does one short peptide do so many protective jobs?
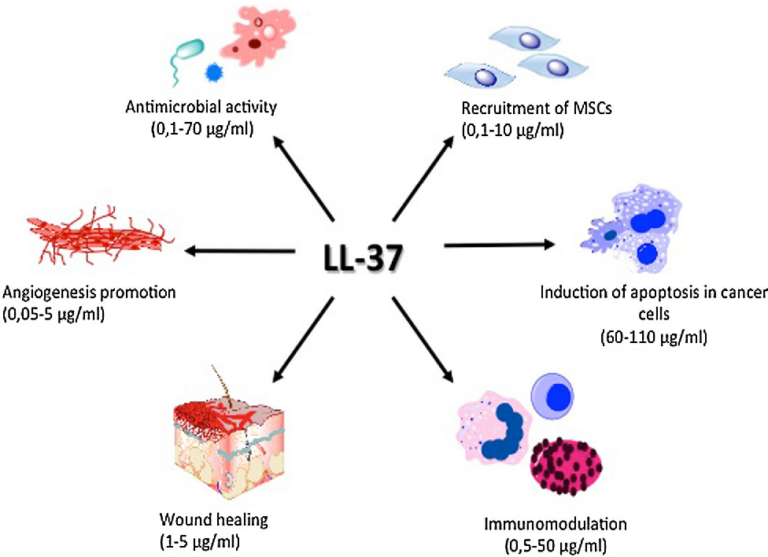
Direct killing is only the headline. LL-37 binds and neutralises lipopolysaccharide (LPS), the endotoxin of Gram-negative bacteria, blunting the inflammatory cascade that LPS would otherwise ignite.1 It is also a chemoattractant, recruiting neutrophils, monocytes and T cells to sites of injury and infection, partly through formyl-peptide receptor signalling.5 And it participates in wound healing, where studies have observed it promoting keratinocyte migration and re-epithelialisation and influencing angiogenesis.5
37 amino acids — the entire functional length of the only cathelicidin humans make
This is the protective face that explains the research interest: a single endogenous molecule that kills pathogens, defuses their toxins, organises the immune response and helps tissue close. In models of impaired healing, low cathelicidin expression has been associated with poor outcomes5, which is part of why the wound-repair literature keeps returning to it.
Where does the same peptide become the problem?
Here the story darkens. LL-37 is strongly cationic, and DNA is strongly anionic; the two form complexes readily.15 In psoriasis, those LL-37–self-DNA complexes are taken up by plasmacytoid dendritic cells and presented to endosomal Toll-like receptor 9, which would normally ignore the host’s own DNA.15 The result is a flood of type-I interferon and a self-sustaining inflammatory loop15 — the peptide effectively converts a fragment of the patient into an alarm signal. LL-37 has even been identified as an autoantigen recognised by T cells in a subset of psoriasis patients11, deepening the case that it is not a bystander but a driver.
The molecule that defuses bacterial DNA can, with the host’s own DNA, arm it.
The same chemistry recurs elsewhere. In rosacea, abnormal processing of cathelicidin generates aberrant peptide forms that correlate with the inflammatory and vascular features of the disease.11 In lupus, LL-37 bound to self-DNA and to neutrophil extracellular traps has been implicated in breaking immune tolerance and sustaining autoantibody production.15 The unifying lesson is uncomfortable: there is no separate “bad” LL-37. It is the same peptide, judged by context, concentration and what it happens to be carrying.
| Face | Mechanism observed | Setting |
|---|---|---|
| Antimicrobial | Membrane disruption of bacteria | In vitro, broad-spectrum |
| Endotoxin control | LPS binding and neutralisation | In vitro / preclinical |
| Immune recruitment | Chemotaxis of leukocytes | Preclinical |
| Repair | Keratinocyte migration, angiogenesis | Cell and animal models |
| Pathology | Self-DNA complexes activate TLR9 / type-I IFN | Psoriasis, lupus |
The same structural property — strong positive charge — underlies both the protective and the pathological columns.
How good is the evidence, honestly?
Thin, in the place that matters most: humans, as a therapy. The protective biology is well characterised mechanistically, but it rests heavily on in-vitro work and animal models, and much of the antimicrobial potency observed in buffer falls off under physiological salt and serum conditions.8 The peptide is also proteolytically fragile, which complicates any attempt to deliver it intact.14 There is no licensed LL-37 medicine; therapeutic candidates — including engineered analogues intended to keep the antimicrobial action while shedding the inflammatory liability — remain preclinical or, at most, early-stage.1
And the double edge is not a footnote — it is the central drug-development obstacle. A molecule that heals at one concentration and inflames at another, depending on what it binds, is exactly the kind of agent regulators and developers approach with caution.2 The honest summary is that LL-37 is a superb piece of basic immunology and an unproven therapeutic.
Where does it sit among other research peptides?
For laboratories building an innate-immunity panel, LL-37 reads naturally alongside other immune-modulating peptides under study. Thymosin Alpha-1, an immune-tuning peptide with a longer clinical history, and ARA-290, an erythropoietin-derived fragment investigated for tissue-protective and anti-inflammatory signalling2, occupy the same conceptual cluster of context-dependent immune modulation. On the repair side, the short tripeptide KPV and the gastric peptide BPC-157 are frequently examined in parallel wound- and inflammation-focused models5, making them common companions in comparative research rather than substitutes. None of these is interchangeable with LL-37; the value of studying them together is precisely the contrast in mechanism.
Condor Research supplies LL-37 strictly as a research-use-only reference material for laboratory investigation, not for human or veterinary use. Because LL-37 is a single defined sequence whose behaviour is exquisitely sensitive to length, charge and truncation, identity and purity documentation is not optional: a current Certificate of Analysis, with HPLC purity and mass-spectrometry confirmation of the exact 37-residue species, is what separates a reproducible experiment from an uninterpretable one. Insist on that paperwork, and treat any peptide lacking it as unknown.
- Humans express a single cathelicidin; its mature, active fragment is the 37-residue, cationic, amphipathic peptide LL-37.
- Its protective repertoire is broad: direct membrane-disrupting antimicrobial action, neutralisation of bacterial LPS, chemotaxis and pro-healing signalling.
- The same peptide is a documented driver of inflammatory pathology — psoriasis, rosacea and lupus — where it shuttles self-DNA into immune cells and breaks tolerance.
- Whether LL-37 heals or harms is context- and concentration-dependent, which is exactly what makes it hard to drug.
- Honest status check: therapeutic application is preclinical or early-stage only; no licensed LL-37 medicine exists, and proteolytic instability is a real obstacle.
- It sits in an innate-immune research cluster alongside Thymosin Alpha-1 and ARA-290, and repair-oriented peptides such as KPV and BPC-157.
Why is LL-37 described as antimicrobial yet implicated in inflammatory disease?
Because both behaviours flow from the same property: a strong positive charge. That charge lets LL-37 disrupt anionic bacterial membranes and neutralise LPS, but it also lets the peptide bind anionic self-DNA. Those complexes can activate intracellular sensors and trigger type-I interferon, driving the inflammation seen in psoriasis and lupus. It is one molecule judged entirely by context.
Is LL-37 an approved drug anywhere?
No. LL-37 is an endogenous human peptide and a heavily studied research molecule, but there is no licensed LL-37 medicine. Therapeutic candidates, including engineered analogues designed to retain antimicrobial action while reducing inflammatory liability, remain preclinical or early-stage. Research-grade LL-37 is a laboratory reference material, not a treatment, and should never be confused with an approved therapy.
What makes LL-37 difficult to develop therapeutically?
Three things. Its antimicrobial potency seen in vitro often weakens under physiological salt and serum. It is proteolytically fragile, complicating delivery of the intact peptide. And its activity is sharply context- and concentration-dependent, so the same molecule can support repair or amplify inflammation. That dual nature is the central obstacle, not a minor caveat, and explains the cautious, preclinical status.
Why does purity documentation matter so much for LL-37 specifically?
Because LL-37's function depends precisely on its 37-residue sequence, charge and helical structure. Truncations, oxidation or sequence errors can alter or abolish activity, and an impure preparation can confound results in unpredictable ways. A current Certificate of Analysis with HPLC purity and mass-spectrometry confirmation of the exact species lets a laboratory know what it is actually testing.